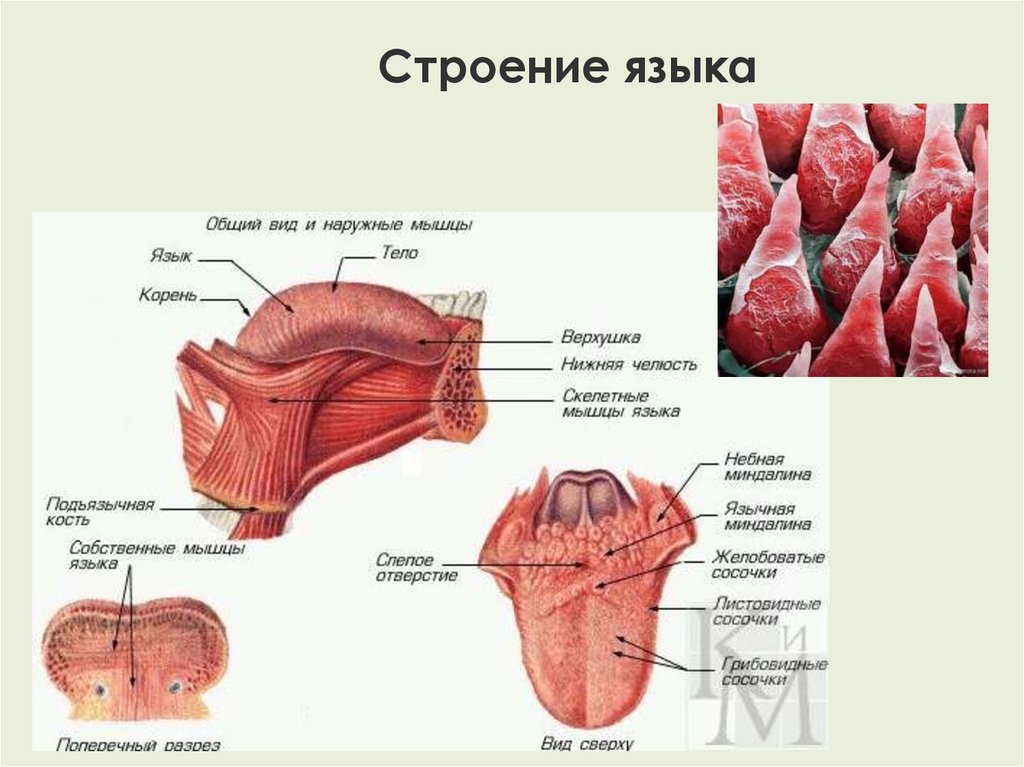
Исследование анатомо-топографических особенностей рисунка слизистой оболочки тыльной поверхности языка в ближайшем и отдаленном постмортальном периоде
Для идентификации личности в качестве одного из возможных критериев предложено использовать рисунок слизистой оболочки спинки языка, анатомическое расположение которого обусловливает доступность его изучения визуальными методами, а также с помощью различных оптических приборов [1].
Более поздними исследованиями [2, 3] был определен комплекс критериев, учитывающий цвет слизистой оболочки языка, расположение, размер и форму сосочков, варианты складчатости. Установлена возрастная и половая динамика. Определено влияние этнических различий внешности на индивидуальные особенности строения языка.
Не вызывает сомнения тот факт, что изменения слизистой оболочки полости рта находятся в тесной взаимосвязи с заболеваниями внутренних органов, поскольку существует тесная анатомо-физиологическая функциональная связь органов желудочно-кишечного тракта с полостью рта. Патологические процессы, развивающиеся в желудке, печени и поджелудочной железе, как правило, приводят к органическим и функциональным нарушениям не только слизистой оболочки полости рта, но и языка.
Патологические процессы, развивающиеся в желудке, печени и поджелудочной железе, как правило, приводят к органическим и функциональным нарушениям не только слизистой оболочки полости рта, но и языка.
Экспертная практика показывает, что нередко объектами исследования судебно-медицинских экспертов являются гнилостно-трансформированные трупы и даже расчлененные их части, что значительно затрудняет решение многих вопросов правоохранительных органов. В связи с изложенным выше перед нами была поставлена задача изучить степень сохранности анатомо-топографических параметров слизистой дорсальной поверхности языка в зависимости от сроков наступления смерти.
Для решения задачи проведено исследование языка у 290 трупов обоего пола. В группу обследуемых вошли 185 трупов мужчин и 105 женщин (в возрасте от 18 до 80 лет). Смерть наступала как от ненасильственных, так и насильственных причин.
Распределение трупного материала по полу и возрасту представлено в таблице.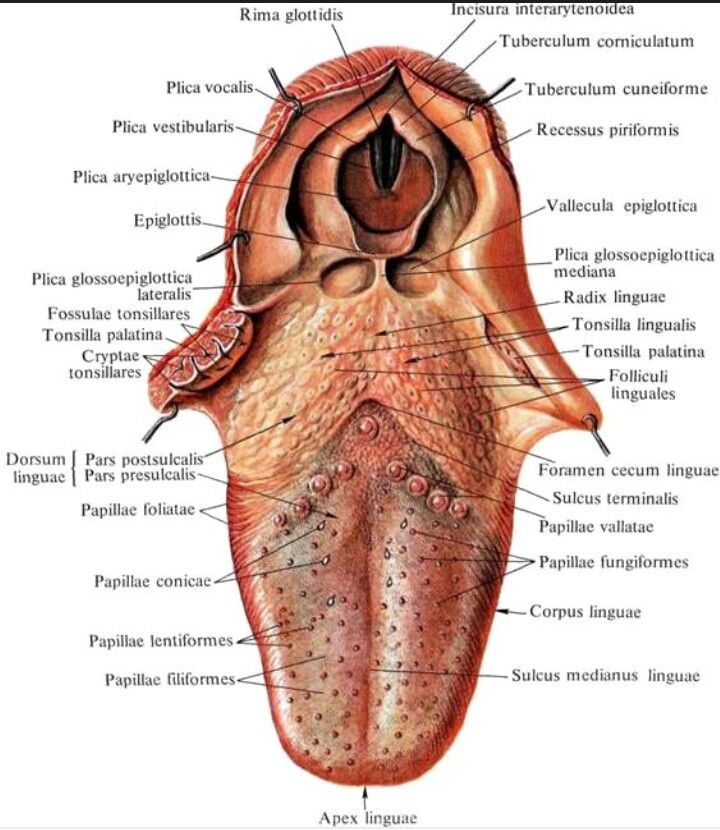
Языки помещали в чашки Петри и хранили в специальном помещении при комнатной температуре (18—20 °С) и влажности воздуха 40—60%. При этом отмечались цвет слизистой оболочки спинки языка, состояние налета, его интенсивность и распространенность, наличие или отсутствие пигментации. Определяли угол терминальной борозды, геометрические размеры, локализацию и количество желобовидных сосочков. Для изучения состояния слизистой оболочки языка в динамике постмортального периода исследовали анатомо-топографические особенности языка с использованием метода визуального осмотра, цифровой фотокамеры SONY (DSC-R1 с разрешением 10,3 megapixels) и графического редактора Adobe Photoshop CS2 для регистрации и анализа изображений в разные сроки после наступления смерти.
При исследовании состояния языка в течение первых 3 дней после наступления смерти слизистая оболочка на всем протяжении была блестящей, бледно-розового цвета (рис. 1).Рисунок 1. Слизистая оболочка дорсальной поверхности языка.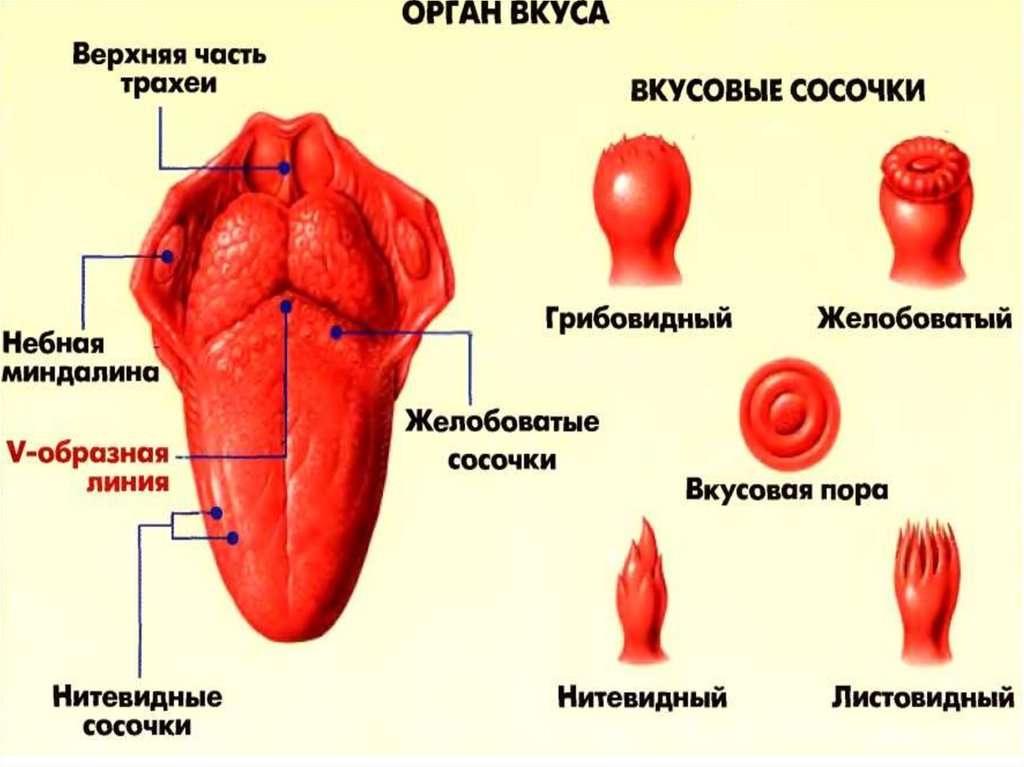
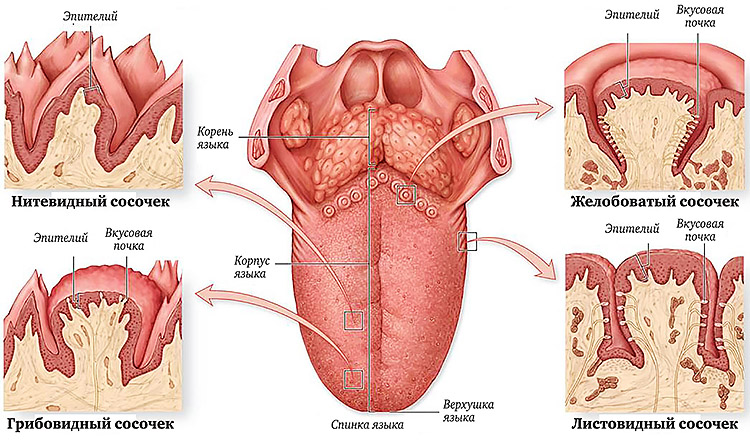
Желобовидные сосочки располагались вдоль терминальной борозды, их количество варьировало, в большинстве случаев было одинаковым на каждой половине языка.
Нитевидные сосочки занимали практически всю поверхность языка. В большинстве случаев они имели коническую форму и заостренные вершины, напоминали выступы слизистой оболочки. Основание сосочков было округлой формы диаметром 0,2—0,3 мм. Встречались также нитевидные сосочки с уплощенными вершинами, напоминающими прямоугольник, овал или треугольник.
В центральной части нитевидные сосочки были более выражены, имели относительно большой размер. На кончике языка сосочки располагались гуще, образуя сплошные массивы, беспорядочно рассредоточенные на незначительном расстоянии друг от друга.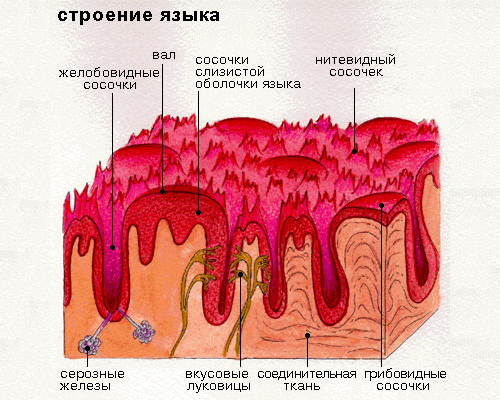
Грибовидные сосочки располагались в основном на кончике и боковой поверхности, а также в области спинки языка и терминальной борозды. На спинке языка их размер был больше, чем на кончике. Сосочки были овальной или округлой формы с четкими границами, возвышались над уровнем нитевидных сосочков. Листовидные сосочки чаще имели форму складок и располагались в боковых отделах задней половины языка.
Такая морфологическая картина в основном сохранялась в течение 7 дней постмортального периода. Через 1 нед после наступления смерти слизистая оболочка языка на всем протяжении была блестящей. Отмечалось изменение цвета слизистой оболочки с бледно-розового на грязно-зеленый с желтоватым оттенком. Складчатость дорсальной поверхности, форма и размер языка практически не изменялись. Количество, цвет и локализацию налета определить было достаточно затруднительно.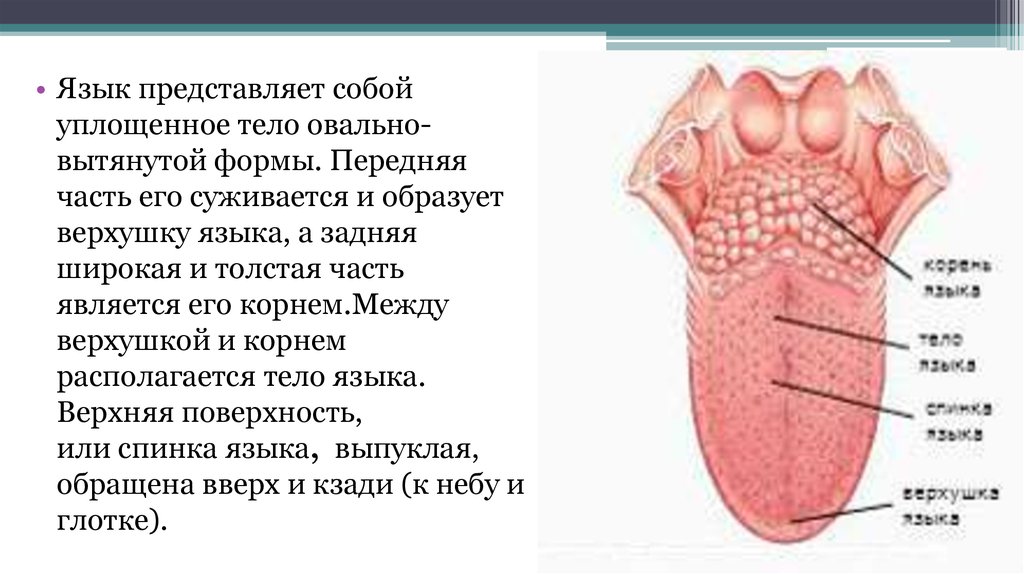
Начиная со 2-й недели после наступления смерти слизистая оболочка языка на всем протяжении имела серовато-коричневый цвет, постепенно превращаясь в грязно-коричневый, выглядела слегка помутневшей. Язык несколько увеличивался в размере, уплощался, а по краям приобретал фестончатость. Четко определялись складки листовидных сосочков, легко можно было определить их количество как справа, так и слева. Грибовидные сосочки выглядели как красные точки разного диаметра. Желобовидные сосочки располагались в виде буквы V, параллельно терминальной борозде.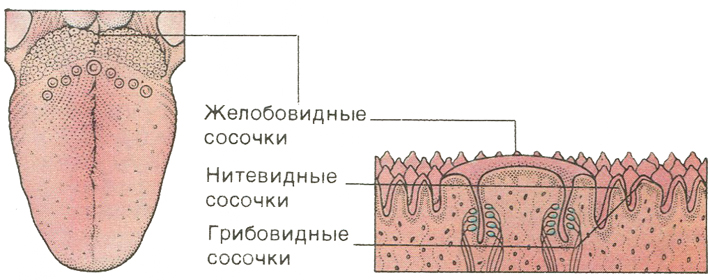
При длительности постмортального периода 3 нед слизистая оболочка языка на всем протяжении приобрела грязно-коричнево-зеленый цвет. Она легко отслаивалась грязно-коричневыми лоскутами с бахромчатыми краями. Вся поверхность была покрыта грязно-серо-желтым слизеподобным налетом, который тянулся нитями при дотрагивании до него пинцетом. Гнилостные пузыри не выявлялись (рис. 2, в).
После удаления налета с поверхности языка в структурных элементах четко просматривались красные точки грибовидных сосочков в местах их обычной локализации. Желобовидные сосочки сохранялись без видимых изменений.
Через 4 нед после наступления смерти слизистая оболочка языка имела значительные гнилостные изменения во всех отделах. Цвет слизистой был грязно-темно-коричневый. На всей поверхности имелись мелкоочаговые дефекты, через которые просматривался мышечный слой в виде распадающейся темно-коричневой массы. Практически все структурные элементы слизистой оболочки дорсальной поверхности языка были практически неразличимы (рис. 2, г).
2, г).
Только желобовидные сосочки в указанный период после наступления смерти оставались без ярко выраженных гнилостных изменений. Без особого усилия можно было определить их количество с обеих сторон. Отмечалось сглаживание валика, окружающего сосочек.
Через 2 мес постмортального периода отмечается резкое усиление гнилостной трансформации языка. Слизистая оболочка со значительными дефектами, грязно-серого цвета с множественными гнилостными пузырями. Мышца языка дряблая, кашицеобразная. Структурные элементы практически не различались, что делало нецелесообразным исследование особенностей языка в более поздние сроки.
Обобщая сказанное выше, можно констатировать, что в сроки до 1 мес после наступления смерти анатомо-топографические особенности слизистой оболочки языка сохраняют свою диагностическую ценность и могут быть использованы для идентификации личности. При этом наиболее значимыми из них являются морфометрические размеры языка, число желобовидных сосочков и угол терминальной борозды, которые сохраняются дольше, чем остальные анатомические элементы языка, это основные времененны`е показатели структурных изменений языка.
Четыре тайны вкуса
Во время приема пищи человеку не составляет труда определить, вкусная ли еда, не пересолена ли она. Но как мы это делаем? Реальна ли карта языка, которую проходят в школе? И какую роль в оценке вкуса играют другие ощущения?
Тайна «пупырышек»
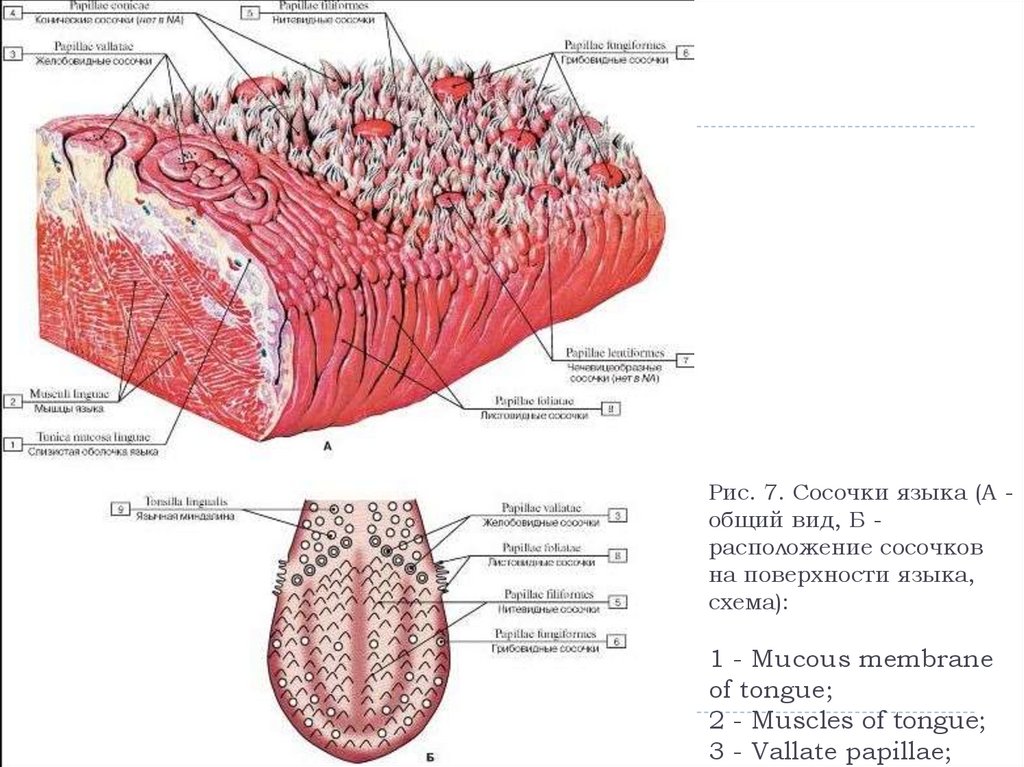
Если вы потрогаете язык, то почувствуете небольшие выпуклости. Они называются вкусовыми сосочками, или папиллами. И на некоторых из них может поместиться более ста вкусовых рецепторов — количество разнится в зависимости от типа сосочков.
|
Нитевидные сосочки, или конические папиллы, расположены в центре языка. Они осязают пищу и удерживают ее во рту. Нитевидные сосочки не содержат вкусовых рецепторов. |
|
|
Грибовидные папиллы распределены по всей поверхности языка, но особенно их много на передней части и по бокам. |
|
|
Желобовидные сосочки расположены около основания языка в форме буквы V, каждый из них по виду напоминает бокал. Они самые крупные среди папилл, но их немного, обычно 7–12. Однако один желобовидный сосочек содержит более 100 рецепторов. |
|
|
На задней части языка по бокам от желобовидных находятся листовидные сосочки. Как и два предыдущих типа, они распознают вкус, но также выполняют тактильную и термическую функции. В каждом листовидном сосочке — более 100 рецепторов. |
Тайна карты языка
Если выделить описанные зоны расположения сосочков, то это напоминает известную по школьным урокам биологии карту языка. Она показывает, на какие вкусы реагируют разные зоны органа. Однако это не совсем так.
У человека в общей сложности от 2 до 9–10 тысяч вкусовых рецепторов. Они рассредоточены на языке, меньше в области мягкого нёба и нёбных дужек, на задней поверхности надгортанника и на внутренней поверхности черпаловидных хрящей. Каждый рецептор содержит комплекс белков, которые способны идентифицировать любой вкус, но могут быть более чувствительны к какому-то одному, однако это различие незначительно. Так, в районе листовидных сосочков больше клеток, реагирующих сильнее на горький вкус, а, например, в передней части языка — на сладкий.
Основными вкусами, которые распознает язык, считаются соленый, горький, кислый и сладкий. Существует еще умами: в переводе с японского — «приятный пикантный вкус». Выделить его смог химик Икэда Кикунаэ в 1908 году. Этот вкус пищевому продукту придает глутамат, а также рибонуклеотиды. Умами можно охарактеризовать как мясной и приятный, он имеет долгое послевкусие.
Комментарий эксперта
Анастасия Орехова, клинический ординатор-неврологАгевзия — медицинский термин, означающий отсутствие вкусовой функции языка.
Агевзия не угрожает жизни, но часто вызывает дискомфорт. Это может привести к потере аппетита, снижению веса, оказать серьезное психологическое воздействие на пациента, в некоторых случаях — потребовать отмены лекарств у уже скомпрометированных (со сниженной или нарушенной иммунной реакцией — примеч. ред.) пациентов.
Существуют состояния, из-за которых люди теряют вкус, например повреждение лицевого и языкоглоточного нервов. Оно чаще всего связано с невритом, вызванным опоясывающим герпесом, рассечением шейных артерий, менингиомой, невриномой или опухолевыми поражениями на основании черепа. Могут страдать от агевзии пациенты с онкологическими заболеваниями в области головы и шеи, получающие лучевую терапию. Нарушения питания, системные состояния, такие как гипотиреоз, сахарный диабет и другие, невралгии и полинейропатии способны изменять вкусовую чувствительность.
Дефицит цинка — причина нарушений восприятия вкуса у здорового человека. Также они могут быть вызваны употреблением наркотиков.
Нередко расстройство вкуса — результат некоторых местных травм (ожогов, рваных ран, хирургического вмешательства) и воспалений, анестезии. На функцию вкуса могут влиять препараты против зубного налета, инфекционные поражения зубов, мягких тканей ротовой полости, полные и частичные съемные протезы, металлические зубные реставрации, дисфункция слюнной железы.
Ряд препаратов способны вызвать нарушение вкусовой чувствительности как побочный симптом. Кроме того, старение или факторы, связанные с ним, делают людей более подверженными дисфункции вкусовой системы. Также, уточнение в связи с последними событиями, потеря вкуса и обоняния — один из симптомов COVID-19.
Как видно, спектр причин агевзии столь широк (от банального несоблюдения гигиены полости рта до генетических заболеваний), что дать какую-либо единую рекомендацию, кроме как вести здоровый образ жизни, достаточно трудно. Во всех случаях при потере или изменении вкусовой чувствительности следует незамедлительно обратиться к врачу, и уже совместно с ним вы обнаружите причину и подберете подходящий вариант лечения и профилактики этого состояния.
Тайна органа вкуса
Вкусовые клетки — отправная точка проводящего пути вкусового анализатора. Они входят в состав вкусовой луковицы, или рецептора. Чтобы ощутить вкус, необходим непосредственный контакт клеток с растворенным веществом. Для этого на вершине луковицы находится пора, выходящая на поверхность сосочка.
Попадая в пору, молекула вещества связывается лишь с соответствующей ей рецепторной клеткой. Так, рецептор сладких веществ — это димер из двух белков T1R2 и T1R3, за умами ответственен димер T1R1/T1R3, а около 30 рецепторов группы T2R позволяют нам чувствовать горечь. Механизм формирования ощущения кислого или соленого отличается от предыдущих, и за первое отвечает повышенная концентрация положительно заряженных ионов водорода, а за второе — солей натрия.
У рецептора есть микроворсинки, которые реагируют на то или иное вещество. Далее возбуждение в виде импульсов передается на чувствительные нейроны. К каждой вкусовой луковице подходят 1–2 нервных волокна.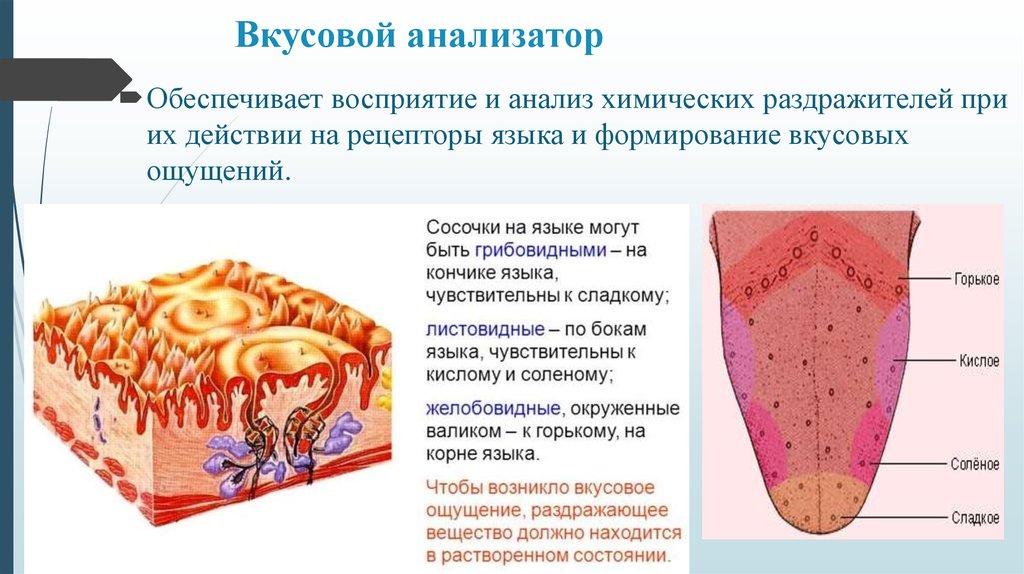 В итоге они собираются в 3 нервных пучка: один проходит по лицевому нерву, другой — по языкоглоточному, а последний — по блуждающему.
В итоге они собираются в 3 нервных пучка: один проходит по лицевому нерву, другой — по языкоглоточному, а последний — по блуждающему.
Сигнал от языка идет по нервам в продолговатый мозг и мост — отделы ствола головного мозга. На их границе находятся первичные вкусовые центры. В них запускаются пищевые (жевание, глотание, выделение желудочного сока) или оборонительные, например рвота, рефлексы. Далее сигнал направляется выше — к гипоталамусу и таламусу. Первый отвечает за эмоциональное восприятие пищи: в частности, именно из-за «фокусов» гипоталамуса беременные женщины могут странно реагировать на привычную им еду и предпочитать необычные вкусовые сочетания. А в медиальных ядрах таламуса происходит идентификация информации, поступающей от нескольких органов чувств, и отправка сигнала в нужную зону мозга.
Конечная точка маршрута — кора больших полушарий. Основной центр вкуса находится на дне боковой борозды в островковой доле. Здесь и возникает полное восприятие вкуса. В 2019 году ученые Колумбийского и Корнелльского университетов (США) и Национального института физиологических наук (Япония) доказали это, используя функциональную магнитно-резонансную томографию и методы статистического анализа.
Тайна других ощущений
В формировании вкуса участвуют не только вкусовые рецепторы, но и осязательные, термические, обонятельные. Ученые Йельского университета определили, что ощущение кислого или соленого появляется при охлаждении краев языка до 20 °С, а сладкого — при согревании краев или кончика до 35 °С. Благодаря тактильным ощущениям мы можем использовать в описании вкуса такие эпитеты, как, например, вяжущий или мучнистый. В носовой полости находятся обонятельные рецепторы, определяющие запах. Сигналы от них и от системы кожной чувствительности поступают в уже упомянутую зону островковой доли, где объединяются с сигналами от вкусовых рецепторов, создавая полную картину ощущения.
Расстояние от языка до островковой доли импульсы проходят примерно за 0,001 секунды. За это ничтожное время мозг успевает определить, какой вкус мы ощущаем. Однако процесс его распознавания хранит еще достаточно тайн, которые ученым только предстоит раскрыть.
Материал опубликован в журнале NewTone
Трехмерные аспекты язычных сосочков и их ядер соединительной ткани в языке крыс: исследование с помощью сканирующего электронного микроскопа
ScientificWorldJournal. 2014; 2014: 841879.
2014; 2014: 841879.
Опубликовано в Интернете 10 ноября 2014 г. doi: 10.1155/2014/841879
, 1 , 1 , 1 , 2 и 2 , 3 , *
Информация об авторе Примечания к статье Информация об авторских правах и лицензии Отказ от ответственности
Целью настоящего исследования было описание трехмерных морфологических характеристик язычных сосочков и их ядер соединительной ткани (CTCs) у крыс Sprague Dawley. Сообщалось о четырех типах папилл на дорсальной поверхности. На поверхности языка располагались нитевидные сосочки, после мацерации эпителия выявлялась коническая и мультифиламентарная форма ЦОК. Сообщалось о грибовидных сосочках в ростральной и средней областях, покрытых плоским эпителием. После удаления эпителия отмечалась форма вулкана с вкусовым отверстием на его вершине. Листовидные сосочки состоят из пяти пар эпителиальных складок, расположенных на латерально-каудальном крае языка. Показано, что после удаления эпителия они ограничены тонкими ламинарными выростами. В каудальной области имелся желобовидный сосочек овальной формы, отграниченный неполной бороздкой. Морфологические характеристики язычных сосочков крыс Sprague Dowley, трехмерные СЭМ-изображения и типы сосочков на дорсальной поверхности были аналогичны описанным ранее у других млекопитающих-грызунов. Методом мацерации выявлены детали внеклеточного матрикса различной формы в виде ядер соединительной ткани.
Показано, что после удаления эпителия они ограничены тонкими ламинарными выростами. В каудальной области имелся желобовидный сосочек овальной формы, отграниченный неполной бороздкой. Морфологические характеристики язычных сосочков крыс Sprague Dowley, трехмерные СЭМ-изображения и типы сосочков на дорсальной поверхности были аналогичны описанным ранее у других млекопитающих-грызунов. Методом мацерации выявлены детали внеклеточного матрикса различной формы в виде ядер соединительной ткани.
Язык заполняет большую часть полости рта и доходит до ротоглотки. Его корень связан с телом и свободной вершиной. На самом деле это мышечный орган, способный к энергичным и точным движениям, таким как удержание, притирание, уход за пищей и манипулирование ею в ротовой полости, а также вокализация [1].
Тело языка состоит из массы переплетенных пучков скелетных поперечно-полосатых мышечных волокон с разнообразным расположением, что обеспечивает большое разнообразие движений языка. Волокна можно разделить на две группы: волокна, которые берут начало вне языка (внешние мышцы), и волокна, которые берут начало внутри языка и вставляются в него. Последние формируют собственные мышцы, изменяющие форму языка [2].
Последние формируют собственные мышцы, изменяющие форму языка [2].
Язык покрыт слизистой оболочкой или, скорее, сплошной слизью разной толщины по всей поверхности. Эпителий толстый и жесткий на дорсальной поверхности, где изнашивание языка больше из-за трения о пищу [3]. Ультраструктурные элементы эпителия различаются по морфологии у разных млекопитающих и содержат несколько типов сосочков. Помимо того, что они связаны с видами животных, вариации могут быть также связаны с типом пищи и адаптацией животного к условиям окружающей среды [4].
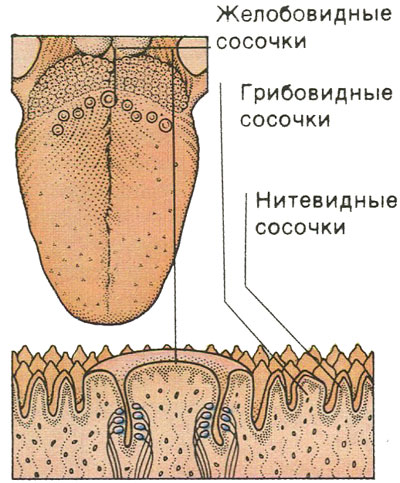
Язычный эпителий состоит из четырех типов сосочков языка, а именно нитевидных, грибовидных, листовидных и желобовидных сосочков, выполняющих механические и вкусовые функции [5]. Однако форма, размер и организация сосочков языка варьируются в зависимости от каждого вида млекопитающих [6].
Нитевидные сосочки густо расположены на всех дорсальных и боковых поверхностях языка. Они помогают манипулировать пищей и увеличивают трение во время жевания. Они преобладают на верхушке и, как правило, наклонены в каудальном отделе, тогда как у корня они видоизменены и имеют различную форму [7]. Различия в морфологии нитевидных сосочков можно наблюдать на верхушке и в средней части языков у некоторых видов. Конические сосочки имеют удлиненную форму с широким основанием и плоской вершиной. Их выстилка состоит из хорошо развитых ороговевших многослойных эпителиальных тканей, основной задачей которых является истирание и защита [8]. На верхушке определяются грибовидные сосочки, отличающиеся большим количеством соединительной ткани и прилежащим эпителиальным слоем. Они хорошо васкуляризированы, но в меньшем количестве, чем нитевидные сосочки, принимают форму «гриба». Они могут представлять собой вкусовые луковицы, обычно расположенные на вершине сосочков, и вовлечены в сенсорную систему, связанную со вкусом [9].].
Они преобладают на верхушке и, как правило, наклонены в каудальном отделе, тогда как у корня они видоизменены и имеют различную форму [7]. Различия в морфологии нитевидных сосочков можно наблюдать на верхушке и в средней части языков у некоторых видов. Конические сосочки имеют удлиненную форму с широким основанием и плоской вершиной. Их выстилка состоит из хорошо развитых ороговевших многослойных эпителиальных тканей, основной задачей которых является истирание и защита [8]. На верхушке определяются грибовидные сосочки, отличающиеся большим количеством соединительной ткани и прилежащим эпителиальным слоем. Они хорошо васкуляризированы, но в меньшем количестве, чем нитевидные сосочки, принимают форму «гриба». Они могут представлять собой вкусовые луковицы, обычно расположенные на вершине сосочков, и вовлечены в сенсорную систему, связанную со вкусом [9].].
Листовидные сосочки, если они присутствуют, расположены на билатеральных краях каудальной области, представляют собой листовидные структуры, отделенные друг от друга инвагинацией слизистой оболочки, и имеют вкусовые почки. Количество листовидных сосочков может варьироваться в зависимости от эволюции каждого вида [10].
Количество листовидных сосочков может варьироваться в зависимости от эволюции каждого вида [10].
Желобовидный сосочек является самым большим из сосочков и расположен в каудальной части дорсальной поверхности, вовлечен в глубокую непрерывную борозду. Количество и форма желобовидных сосочков широко варьируют и зависят от анализируемых видов [11]. Они различаются по размеру и форме, от круглых до плоских, лежащих параллельно или рядами с каждой стороны хвостового отдела. В нижней части желобовидных сосочков обнаруживаются группы желез, также известные как железы фон Эбнера. Это слюнные серозные железы, протоки которых открываются в основании борозд сосочков и выделяют водянистую жидкость, растворяющую пищевое содержимое и облегчающую восприятие вкуса [12]. В отряде Rodentia было проведено несколько исследований на мышах [13, 14], морской свинке [15], агути [12], белке-летяге [16], американском бобре [17] и низменной паке [7]. Однако при изучении язычных сосочков крыс можно отметить различную форму на поверхности и их соединительнотканные ядра (СТС) в образцах, обработанных раствором NaOH. Типы и подтипы сосочков языка млекопитающих грызунов могут иметь несколько различий в морфологических характеристиках классификации, форме и вкусовых рецепторах. Целью настоящего исследования было описание трехмерных морфологических характеристик язычных сосочков и их ядер соединительной ткани (CTC) крыс Sprague Dawley с использованием методов сканирующего электронного микроскопа.
Типы и подтипы сосочков языка млекопитающих грызунов могут иметь несколько различий в морфологических характеристиках классификации, форме и вкусовых рецепторах. Целью настоящего исследования было описание трехмерных морфологических характеристик язычных сосочков и их ядер соединительной ткани (CTC) крыс Sprague Dawley с использованием методов сканирующего электронного микроскопа.
Были исследованы четыре языка четырехмесячных самцов крыс, вид, отряд: Rodentia. Животных содержали в полипропиленовых клетках с водой и рационом «вволю», в 12-часовом световом/темновом периоде при средней температуре 25 ± 2°С. Это исследование было одобрено Комитетом по этике использования животных (CEUA) Института биомедицинских исследований Университета Сан-Паулу, Бразилия (159/2010).
2.1. Сканирующая электронная микроскопия
Языки ( n = 4) погружали в раствор модифицированного фиксатора Карновского по методу Ватанабэ и Ямада [18]. Затем образцы промывали в буферном растворе и разделяли по разным методикам. Традиционная методика включала промывание образцов дистиллированной водой в течение 2 часов при комнатной температуре для анализа поверхности эпителия. Для мацерации остальные образцы промывали дистиллированной водой и погружали в 10% водный раствор гидроксида натрия (NaOH) на 4 дня при комнатной температуре [19].–21] для удаления эпителиальной поверхности и анализа соединительнотканного ядра (СТС). Затем их промывали в дистиллированной воде с частыми заменами в течение двух дней при температуре 4°С. После этого этапа все образцы были обезвожены в возрастающей серии спиртов и высушены в сушилке для критической точки (Balzers CPD-030) с жидким СО2. Образцы устанавливали на металлическую основу, покрытую ионами золота (Balzers-040 SDC) [22], и исследовали с помощью сканирующего электронного микроскопа LEO 435 VP.
Традиционная методика включала промывание образцов дистиллированной водой в течение 2 часов при комнатной температуре для анализа поверхности эпителия. Для мацерации остальные образцы промывали дистиллированной водой и погружали в 10% водный раствор гидроксида натрия (NaOH) на 4 дня при комнатной температуре [19].–21] для удаления эпителиальной поверхности и анализа соединительнотканного ядра (СТС). Затем их промывали в дистиллированной воде с частыми заменами в течение двух дней при температуре 4°С. После этого этапа все образцы были обезвожены в возрастающей серии спиртов и высушены в сушилке для критической точки (Balzers CPD-030) с жидким СО2. Образцы устанавливали на металлическую основу, покрытую ионами золота (Balzers-040 SDC) [22], и исследовали с помощью сканирующего электронного микроскопа LEO 435 VP.
Анализы с помощью сканирующего электронного микроскопа показали, что нитевидные сосочки были наиболее многочисленны на дорсальной поверхности языка с меньшим количеством на каудальной области. Они имеют форму и диаметр в зависимости от региона. В ростральной области сосочки низкие конусовидные, с закругленной вершиной (). Удлиненные и многофиламентные сосочки могут быть обнаружены в средней и каудальной областях с соответствующими длинными и тонкими нитями (1). После удаления эпителия в ЦОК нитевидных сосочков (ростральная область) выявляются невысокие, с конусовидной вершиной, основанием шире вершины и тонкими пучками в межсосочковой зоне (), отличающиеся от других нитевидных сосочков. При большем увеличении обнаружено от трех до четырех филаментов (многофиламентных), начиная с третьей средне-верхней части ЦОК (). Грибовидные сосочки располагались в ростральной и средней областях дорсальной поверхности. Они очень многочисленны в ростральной области, имеют округлую форму и куполообразную вершину. В деталях можно отметить, что поверхность сосочков состоит из плоского эпителия и вкусового отверстия на вершине поверхности (). После удаления эпителия выявлялась форма вулкана и наблюдалась полость для вкусовой почки на верхушке сосочков ().
Они имеют форму и диаметр в зависимости от региона. В ростральной области сосочки низкие конусовидные, с закругленной вершиной (). Удлиненные и многофиламентные сосочки могут быть обнаружены в средней и каудальной областях с соответствующими длинными и тонкими нитями (1). После удаления эпителия в ЦОК нитевидных сосочков (ростральная область) выявляются невысокие, с конусовидной вершиной, основанием шире вершины и тонкими пучками в межсосочковой зоне (), отличающиеся от других нитевидных сосочков. При большем увеличении обнаружено от трех до четырех филаментов (многофиламентных), начиная с третьей средне-верхней части ЦОК (). Грибовидные сосочки располагались в ростральной и средней областях дорсальной поверхности. Они очень многочисленны в ростральной области, имеют округлую форму и куполообразную вершину. В деталях можно отметить, что поверхность сосочков состоит из плоского эпителия и вкусового отверстия на вершине поверхности (). После удаления эпителия выявлялась форма вулкана и наблюдалась полость для вкусовой почки на верхушке сосочков ().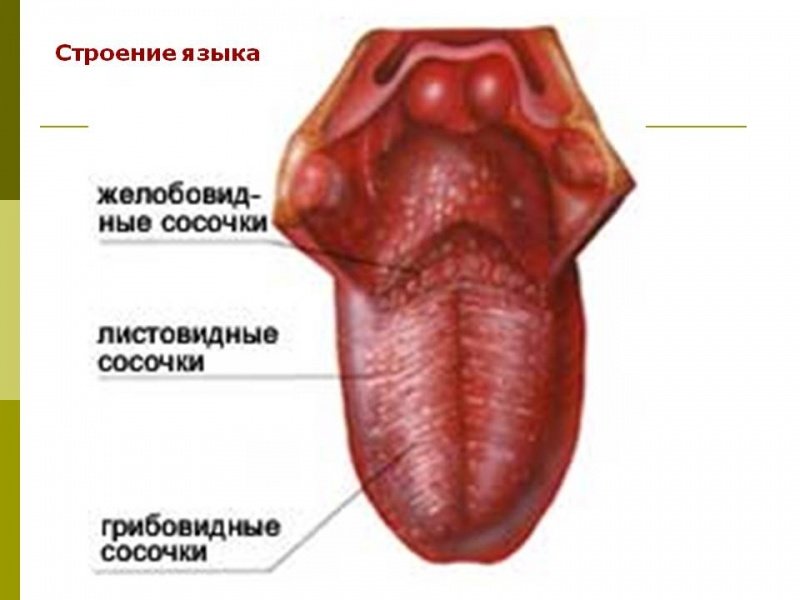
Открыть в отдельном окне
Сканирующие электронные микрофотографии — нитевидные и грибовидные сосочки крыс. (а) Коническая нитевидная форма (стрелки) и (б) после мацерации с NaOH ЦОК нитевидных сосочков представлены коническими (стрелки) и пучками (*) в межпапиллярной зоне. (в) Многофиламентные сосочки (стрелки) и (г) их ЦОК выявили от трех до четырех филаментов (стрелки) в третьей верхней части. (e) На поверхности видны грибовидные сосочки (стрелка) куполообразной формы между коническими нитевидными сосочками и выделенным вкусовым отверстием (стрелка). (f) После удаления эпителия ЦОК грибовидных сосочков приобрели форму вулкана с полостью наверху для вкусовых рецепторов (стрелка). Бары: 30 мк м (а, б, е), 100 мк м (в, д), 10 мк м (г).
Листовидные сосочки представлены пятью парами эпителиальных складок, разделенных параллельными бороздками на латерально-каудальном крае языка (). После удаления эпителия наблюдались ЦОК с широкими овальными бороздками, ограниченными ламинарными выступами (). Желобовидный сосочек расположен в середине каудального отдела, овальной формы, обернутый неполной бороздкой на задней части (). При большем увеличении видно, что сосочки выстланы плоским эпителием (1). После удаления эпителия наблюдали строение ЦОК в виде толстого пучка, окаймленного широкой бороздкой. На боковых стенках видны отверстия слюнных протоков (рис. и ).
Желобовидный сосочек расположен в середине каудального отдела, овальной формы, обернутый неполной бороздкой на задней части (). При большем увеличении видно, что сосочки выстланы плоским эпителием (1). После удаления эпителия наблюдали строение ЦОК в виде толстого пучка, окаймленного широкой бороздкой. На боковых стенках видны отверстия слюнных протоков (рис. и ).
Открыть в отдельном окне
Сканирующие электронные микрофотографии — листовидные и желобовидные сосочки крыс. (а) Листовидные сосочки образованы складками эпителия (стрелки) и разделены параллельными бороздками. (б) После удаления эпителиальной ткани ЦОК листовидных сосочков имели широкие борозды, ограниченные ламинарными выступами (стрелки). (в) желобовидный сосочек (стрелка) расположен в каудальной области и (г) после удаления эпителия общие аспекты ЦОК (*), отграниченные ЦОК нитевидных сосочков и протоков слюнных желез (стрелки), наблюдались. (e) При большем увеличении был отмечен плоский эпителий (стрелки) на поверхности.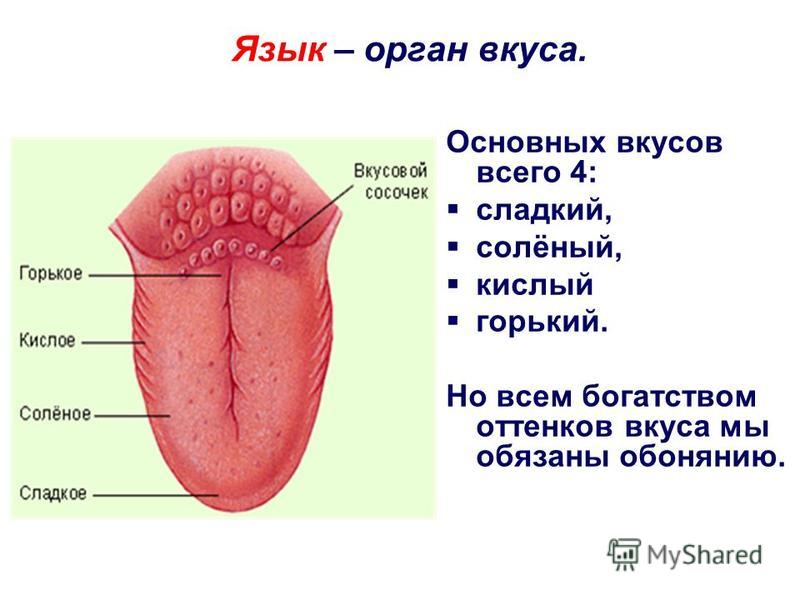 (f) При большем увеличении было обнаружено строение ЦОК в виде толстого пучка (*), окаймленного широкой бороздкой (стрелка). Бары: 100 мк м (а, б, д, е) и 300 мк м (в, г).
(f) При большем увеличении было обнаружено строение ЦОК в виде толстого пучка (*), окаймленного широкой бороздкой (стрелка). Бары: 100 мк м (а, б, д, е) и 300 мк м (в, г).
Результаты, полученные в этом исследовании, выявили в основном трехмерные характеристики сосочков языка и исходное расположение ядер соединительной ткани (CTC) у крыс линии Sprague Dawley. В этих данных на дорсальной поверхности лингвального эпителиального слоя наблюдались четыре типа сосочков: нитевидные, грибовидные, листовидные и желобчатые, подобные тем, о которых сообщают Ciena et al. [12].
Нитевидные сосочки имели низкую конусообразную форму в ростральной области, хотя сосочки были удлиненными и многонитевыми в средней и каудальной областях. После удаления эпителия ЦОК нитевидных сосочков были низкими, с конусовидной вершиной в ростральной области. Однако в мультифиламентарных сосочках выявляются три-четыре нити в средней и каудальной областях. Каждая цилиндрическая нить лежала на верхнем отделе, начиная с середины трети соединительного сосочка.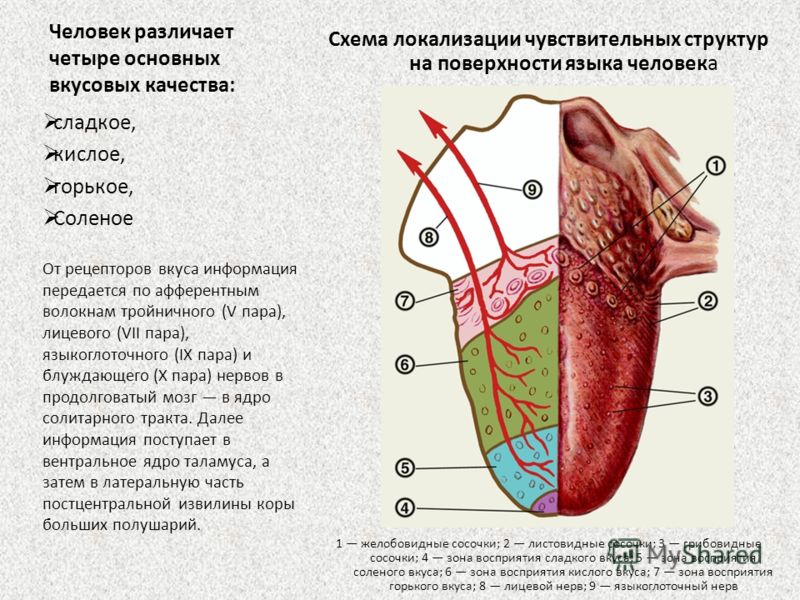 Согласно Kilinc et al. [9], нитевидные сосочки наиболее многочисленны на языке млекопитающих грызунов, распределены по всей дорсальной поверхности эпителия и уменьшены в каудальной области [12]. Морфологические характеристики нитевидных сосочков крыс отличались от таковых у других грызунов, как это было сообщено для белки-летяги. Эмура и др. [16] сообщили, что последний вид представлен коническими сосочками в каудальной области и удлиненными сосочками в ростральной области. Эти морфологические характеристики были аналогичны нитевидным сосочкам, описанным на языке морской свинки, как описано Watanabe et al. [15], а на языке патагонской морской свинки, как описано Emura et al. [23].
Согласно Kilinc et al. [9], нитевидные сосочки наиболее многочисленны на языке млекопитающих грызунов, распределены по всей дорсальной поверхности эпителия и уменьшены в каудальной области [12]. Морфологические характеристики нитевидных сосочков крыс отличались от таковых у других грызунов, как это было сообщено для белки-летяги. Эмура и др. [16] сообщили, что последний вид представлен коническими сосочками в каудальной области и удлиненными сосочками в ростральной области. Эти морфологические характеристики были аналогичны нитевидным сосочкам, описанным на языке морской свинки, как описано Watanabe et al. [15], а на языке патагонской морской свинки, как описано Emura et al. [23].
Грибовидные сосочки были распределены в ростральной и средней областях дорсальной поверхности. Отчеты были аналогичны описаниям на языке других грызунов, таких как агути [12], слепой землекоп [9] и пака [7]. После удаления эпителия была обнаружена форма вулкана, а на вершине сосочков была видна полость, вероятное местонахождение вкусовых рецепторов, подобных тем, которые описаны на языке агути [12]. Современная литература показывает, что для описания различных форм грибовидных сосочков используются другие термины, такие как «цветочный бутон» у патагонской морской свинки [23] и «кулак» у американской клещевины [17] и морской свинки [24].
Современная литература показывает, что для описания различных форм грибовидных сосочков используются другие термины, такие как «цветочный бутон» у патагонской морской свинки [23] и «кулак» у американской клещевины [17] и морской свинки [24].
В листовидных сосочках выявлено пять складок эпителия, разделенных параллельными бороздками на латерально-каудальном крае языка. После удаления эпителия наблюдались широкие овальные бороздки, ограниченные ламинарными выростами. Локализация таких сосочков аналогична таковой у паки [7], белки-летяги [16] и морской свинки [15]. По данным Emura et al. [23], листовидные сосочки у грызунов хорошо развиты. Кроме того, количество эпителиальных складок может варьироваться в зависимости от каждого вида. На языке агути было 12 пар хорошо сконструированных лиственных сосочков [12] и от 22 до 25 пар на языке американской клещевины [17].
В центрально-каудальной области обнаружен желобовидный сосочек, ограниченный неполной бороздкой в верхней части.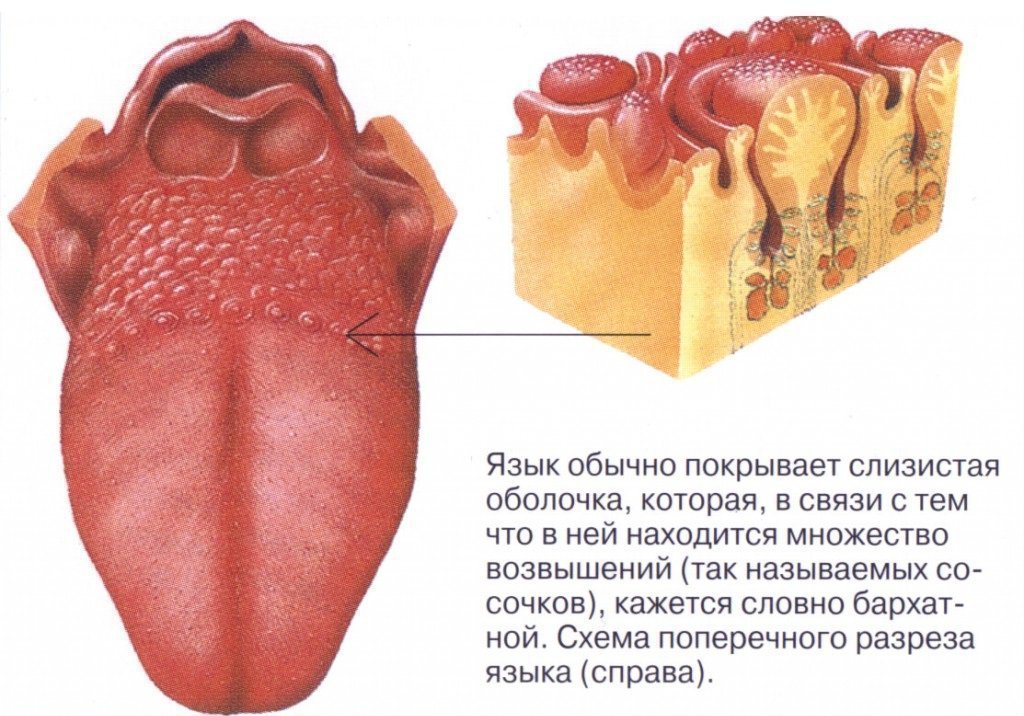 После удаления эпителия отмечалось образование толстого пучка, окаймленного широкой бороздкой ЦОК. Количество и форма этих сосочков различались в зависимости от вида. Количество папилл у рыжей полевки [25] такое же, как у крысы, только с одним папиллом. Кроме того, у паки [7], патагонской морской свинки [23] и слепыша [9] имеется пара параллельных папилл, тогда как у белки-летяги три папиллы [16]. Однако у агути было зарегистрировано четыре сосочка, наибольшее количество у грызунов, как описано Ciena et al. [12].
После удаления эпителия отмечалось образование толстого пучка, окаймленного широкой бороздкой ЦОК. Количество и форма этих сосочков различались в зависимости от вида. Количество папилл у рыжей полевки [25] такое же, как у крысы, только с одним папиллом. Кроме того, у паки [7], патагонской морской свинки [23] и слепыша [9] имеется пара параллельных папилл, тогда как у белки-летяги три папиллы [16]. Однако у агути было зарегистрировано четыре сосочка, наибольшее количество у грызунов, как описано Ciena et al. [12].
Морфологические характеристики язычных сосочков крыс Sprague Dawley, трехмерные СЭМ-изображения и типы сосочков на дорсальной поверхности были аналогичны описанным ранее у других млекопитающих-грызунов. Методом мацерации выявлены детали внеклеточного матрикса различной формы в виде ядер соединительной ткани.
Авторы заявляют об отсутствии конфликта интересов в отношении публикации данной статьи.
1. Дайс К. М., Сак В. О., Венсинг К. Дж. Г. Ветеринарная анатомия .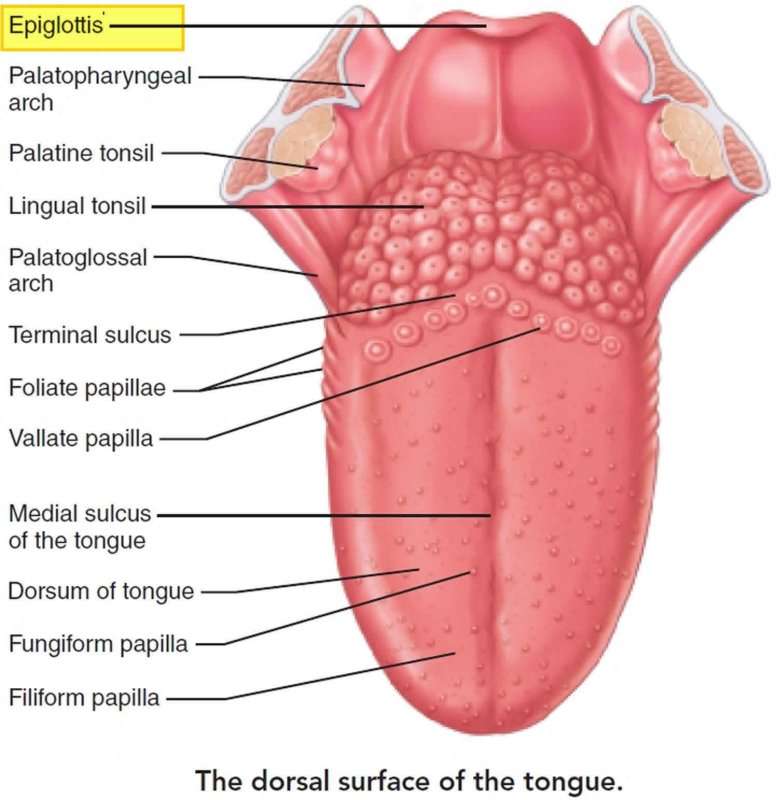 4-й. Рио-де-Жанейро, Бразилия: Elsevier; 2010. [Google Scholar]
4-й. Рио-де-Жанейро, Бразилия: Elsevier; 2010. [Google Scholar]
2. Gartner P.L., Hiatt L.J. Tratado de Histologia em Cores . 3-й. Рио-де-Жанейро, Бразилия: Elsevier; 2007. [Google Scholar]
3. Ferreira R. J., Carvalho A. E., Souza W., Alvarenga F. B., Rodrigues F. B. Anatomia da Artéria Lingual Profunda em Sus scrofa domestica, LINNAEUS, 1758. Ciência Animal Brasileira . 2011;12(2):298–305. [Академия Google]
4. Miyawaki Y., Yoshimura K., Shindo J., Kageyama I. Световое и сканирующее электронное микроскопическое исследование языка и язычных сосочков обыкновенного енота, Procyon lotor . Окаджима Folia Anatomica Japonica . 2010;87(2):65–73. doi: 10.2535/ofaj.87.65. [PubMed] [CrossRef] [Google Scholar]
. Pesquisa Veterinária Brasileira . 2011;31(1):67–73. [Google Scholar]
6. Zheng J., Kobayashi K. Сравнительное морфологическое исследование язычных сосочков и их соединительнотканных ядер (CTC) у мунтжака рифа ( Muntiacus reevesi ) Annals of Anatomy . 2006;188(6):555–564. doi: 10.1016/j.aanat.2006.05.014. [PubMed] [CrossRef] [Google Scholar]
2006;188(6):555–564. doi: 10.1016/j.aanat.2006.05.014. [PubMed] [CrossRef] [Google Scholar]
7. Массоли Б.К.М., Рибейро К.Р.П., Виейра Г.Л., Менезес Т.Л., Лима О.М., Соуза Р.Р., Мачадо Ф.Р.М., Сантос К.Л.А.0045 Cuniculus paca (Rodentia: Cuniculidae) Revista Biotemas . 2013;26(4):167–177. [Google Scholar]
8. Кобаяши К., Кумакура М., Йошимура К., Нонака К., Мураяма Т., Хеннеберг М. Сравнительное морфологическое исследование язычных сосочков и их соединительнотканных стержней коалы. Анатомия и эмбриология . 2003;206(4):247–254. [PubMed] [Google Scholar]
9. Kilinc M., Erdogan S., Ketani S., Ketani M.A. Морфологическое исследование с помощью сканирующей электронной микроскопии язычных сосочков ближневосточного слепыша ( Spalax ehrenbergi , Nehring, 1898) Journal of Veterinary Medicine Series C: Anatomia Histologia Embryologia . 2010;39(6):509–515. doi: 10.1111/j.1439-0264.2010.01022.x. [PubMed] [CrossRef] [Google Scholar]
10. Эмура С., Хаякава Д., Чен Х., Шомура С. Морфология дорсальных язычных сосочков у японской макаки и саванной обезьяны. Анатомия, гистология, эмбриология . 2002;31(5):313–316. doi: 10.1046/j.1439-0264.2002.00378.x. [PubMed] [CrossRef] [Академия Google]
Эмура С., Хаякава Д., Чен Х., Шомура С. Морфология дорсальных язычных сосочков у японской макаки и саванной обезьяны. Анатомия, гистология, эмбриология . 2002;31(5):313–316. doi: 10.1046/j.1439-0264.2002.00378.x. [PubMed] [CrossRef] [Академия Google]
11. Эль Шараби А. А., Эль-Генди С. А., Альсафи М. А., Номир А. Г., Вакисака С. Морфологические вариации желобовидных сосочков у некоторых видов млекопитающих. Международная анатомическая наука . 2014; 89: 161–170. doi: 10.1007/s12565-013-0215-9. [PubMed] [CrossRef] [Google Scholar]
12. Ciena A.P., de Sousa Bolina C., de Almeida S.R.Y., et al. Структурные и ультраструктурные особенности языка агути ( Dasyprocta aguti Linnaeus, 1766) Journal of Anatomy . 2013;223(2):152–158. doi: 10.1111/joa.12065. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
13. Kim J.-Y., Mochizuki T., Akita K., Jung H.-S. Морфологические доказательства важности эпителиальной ткани при развитии языка у мышей. Экспериментальные исследования клеток . 2003;290(2):217–226. doi: 10.1016/S0014-4827(03)00319-7. [PubMed] [CrossRef] [Google Scholar]
Экспериментальные исследования клеток . 2003;290(2):217–226. doi: 10.1016/S0014-4827(03)00319-7. [PubMed] [CrossRef] [Google Scholar]
14. Абэ М., Осава Т. Строение интерстициальных поверхностей эпителиальных базальных мембран слизистой оболочки полости рта, десен и языка мышей. Архив биологии полости рта . 1999;44(7):587–594. doi: 10.1016/S0003-9969(99)00029-1. [PubMed] [CrossRef] [Google Scholar]
15. Ватанабе И.-С., Дос Сантос Хеммерле К. А., Диас Ф. Дж., Кури Д. П., да Силва М. С. П., Состинес М. К., Дос Сантос Т. С., Гимарайнш Ж. П., Миглино М. А. Структурная характеристика языка водосвинки ( Hydrochaeris hydrochaeris ) с помощью световой, сканирующей и просвечивающей электронной микроскопии. Исследования и техника в области микроскопии . 2013;76(2):141–155. doi: 10.1002/jemt.22145. [PubMed] [CrossRef] [Академия Google]
16. Эмура С., Тамада А., Хаякава Д., Чен Х., Джамали М., Тагучи Х., Шомура С. СЭМ-исследование дорсальной язычной поверхности белки-летяги, Petaurista leucogenys
. Анналы анатомии . 1999;181(5):495–498. doi: 10.1016/S0940-9602(99)80033-8. [PubMed] [CrossRef] [Google Scholar]
Анналы анатомии . 1999;181(5):495–498. doi: 10.1016/S0940-9602(99)80033-8. [PubMed] [CrossRef] [Google Scholar]
17. Shindo J., Yoshimura K., Kobayashi K. Сравнительное морфологическое исследование стереоструктуры язычных сосочков и их соединительнотканных ядер американского бобра ( Castor canadensis ) Okajimas Folia Anatomica Japonica . 2006;82(4):128–137. [PubMed] [Google Scholar]
18. Ватанабэ И., Ямада Э. Тонкая структура пластинчатых нервных окончаний, обнаруженных в деснах крыс. Archivum Histologicum Japonicum . 1983;46(2):173–182. [PubMed] [Google Scholar]
19. Отани О. Трехмерная организация волокон соединительной ткани поджелудочной железы человека: сканирующее электронное микроскопическое исследование тканей, обработанных NaOH. Archivum Histologicum Japonicum . 1987; 50 (5): 557–566. [PubMed] [Google Scholar]
20. де Соуза Болина С., де Соуза Болина-Матос Р., де Матос Алвес П. Х., Пульзато Кури Д. , Сиена А. П., Ватанабэ И.-С. Трехмерные аспекты структурных характеристик и ангиоархитектоники почек взрослых и старых крыс Wistar: исследование с помощью сканирующей электронной микроскопии. Исследования и техника в области микроскопии . 2013;76(5):538–544. doi: 10.1002/jemt.22197. [PubMed] [CrossRef] [Академия Google]
, Сиена А. П., Ватанабэ И.-С. Трехмерные аспекты структурных характеристик и ангиоархитектоники почек взрослых и старых крыс Wistar: исследование с помощью сканирующей электронной микроскопии. Исследования и техника в области микроскопии . 2013;76(5):538–544. doi: 10.1002/jemt.22197. [PubMed] [CrossRef] [Академия Google]
21. Benetti E.J., Pícoli L.C., Guimarães J.P., Motoyama A.A., Miglino M.A., Watanabe L.-S. Характеристика нитевидных, грибовидных и желобовидных сосочков и поверхности интерфейса эпителий-соединительная ткань слизистой оболочки языка гривистого ленивца ( Bradypus torquatus , Iliger, 1811): световая и сканирующая электронная микроскопия. Журнал ветеринарной медицины, серия C: Anatomia Histologia Embryologia . 2009;38(1):42–48. doi: 10.1111/j.1439-0264.2008.00890.x. [PubMed] [CrossRef] [Академия Google]
22. Duro C.C., Ciena A.P., de Almeida S.R.Y., et al. Качественное исследование височно-нижнечелюстной синовиальной оболочки молодых, взрослых и старых крыс Wistar с использованием световой, сканирующей и просвечивающей электронной микроскопии.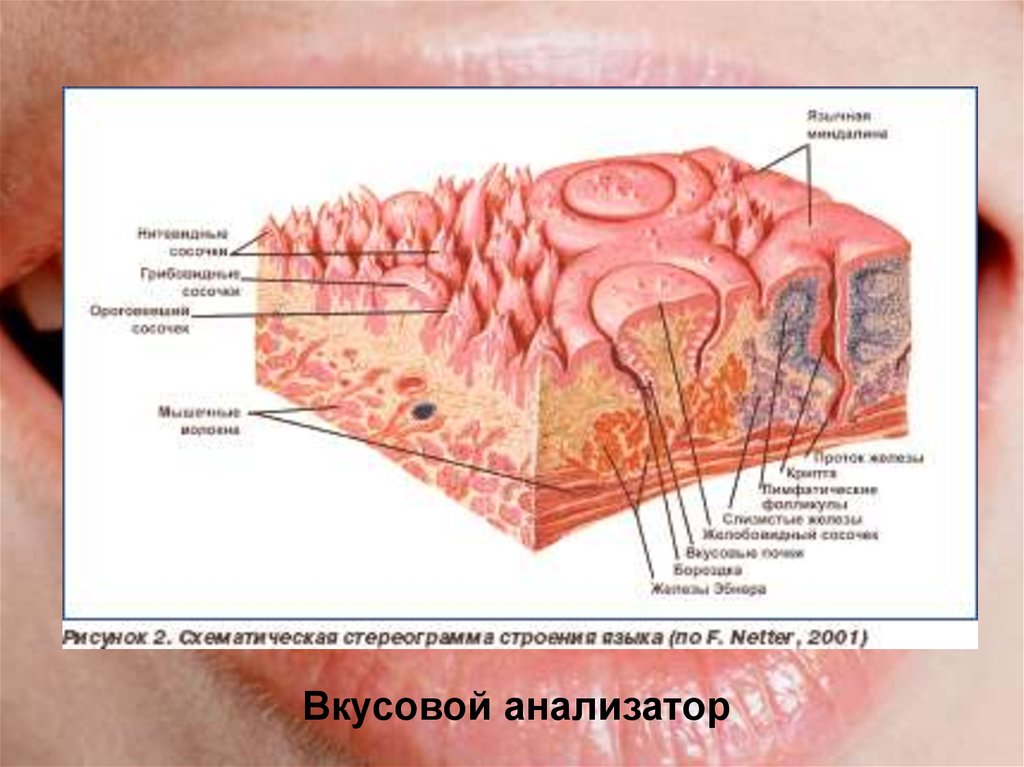 Исследования и техника в области микроскопии . 2012;75(11):1522–1527. doi: 10.1002/jemt.22095. [PubMed] [CrossRef] [Google Scholar]
Исследования и техника в области микроскопии . 2012;75(11):1522–1527. doi: 10.1002/jemt.22095. [PubMed] [CrossRef] [Google Scholar]
23. Эмура С., Окумура Т., Чен Х. Морфология язычных сосочков у патагонской морской свинки. Окадзимас Folia Anatomica Japonica . 2011;88(3):121–125. doi: 10.2535/ofaj.88.121. [PubMed] [CrossRef] [Google Scholar]
24. Кобаяши К. Трехмерная архитектура соединительнотканной сердцевины язычных сосочков морской свинки. Анатомия и эмбриология . 1990;182(3):205–213. [PubMed] [Google Scholar]
25. Jackowiak H., Godynicki S. Распределение и строение язычных сосочков на языке рыжей полевки Clethrionomys glareolus . Фолиа Морфология . 2005;64(4):326–333. [PubMed] [Google Scholar]
вкус%2520анатомия
Вкусовая система состоит из 3-х типов вкусовых сосочков, на которых расположены вкусовые луковицы. Грибовидные сосочки, представляющие собой грибовидные структуры, расположены ближе к передней части языка. Каждый грибовидный сосочек обычно содержит 3-5 вкусовых луковиц. Округлые сосочки расположены ближе к задней части языка, и в отличие от грибовидных сосочков, каждая из них содержит более 100 вкусовых рецепторов. Выступы и бороздки, расположенные по бокам языка, представляют собой листовидные сосочки. Подобно желобчатым сосочкам, листовидные сосочки также содержат более 100 вкусовых луковиц каждый. Сосочки четвертого типа, нитевидные, также существуют, но не содержат вкусовых рецепторов.
Каждый грибовидный сосочек обычно содержит 3-5 вкусовых луковиц. Округлые сосочки расположены ближе к задней части языка, и в отличие от грибовидных сосочков, каждая из них содержит более 100 вкусовых рецепторов. Выступы и бороздки, расположенные по бокам языка, представляют собой листовидные сосочки. Подобно желобчатым сосочкам, листовидные сосочки также содержат более 100 вкусовых луковиц каждый. Сосочки четвертого типа, нитевидные, также существуют, но не содержат вкусовых рецепторов.
Каждая вкусовая почка состоит из 30-100 вкусовых рецепторных клеток. Клетки вкусовых рецепторов представляют собой длинные тонкие клетки, ориентированные перпендикулярно поверхности языка. Каждая вкусовая рецепторная клетка одним концом обращена в полость рта и имеет микроворсинки на своей поверхности для усиления контакта со раздражителями. Противоположный конец клетки вкусового рецептора контактирует с нервными волокнами, которые впадают в языкоглоточный нерв, барабанную струну или блуждающий нерв, в зависимости от расположения вкусовой почки.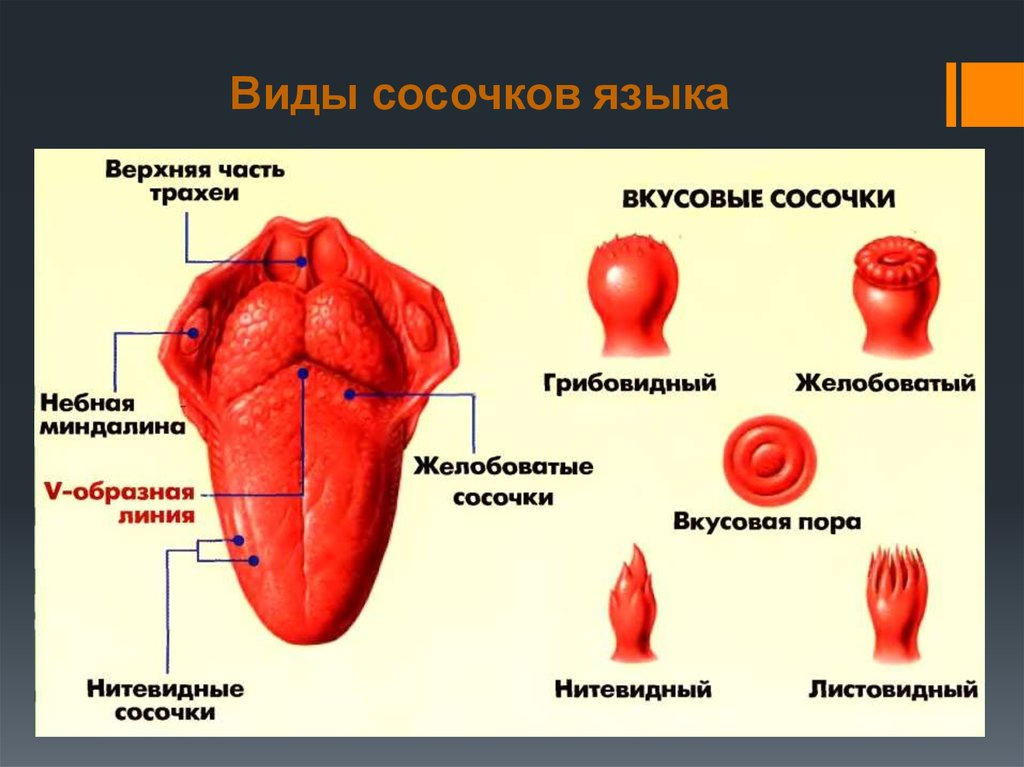 1
1
Восприятие вкуса
Когда раздражители попадают в полость рта, они могут связываться с рецепторами мембраны вкусовых клеток, проходить через определенные каналы или активировать ионные каналы. Затем эти процессы заставляют вкусовые клетки высвобождать нейротрансмиттеры, посылая сигнал в мозг.
Способ, которым различные типы стимулов вызывают вкусовые реакции, до сих пор полностью не изучен. Считается, что сладкий и горький вкус действуют посредством специфических рецепторов, связанных с G-белком, T1R и T2R соответственно. Было показано, что T1R GPCR для сладкого вкуса имеет несколько сайтов связывания, используемых сахарами, искусственными подсластителями и антагонистами сладкого вкуса. Горький вкус может быть вызван гораздо большим числом и более разнообразным набором соединений, чем сладкий вкус. Большое разнообразие горьких соединений указывает на то, что ни один рецептор не может реагировать на все горькие соединения.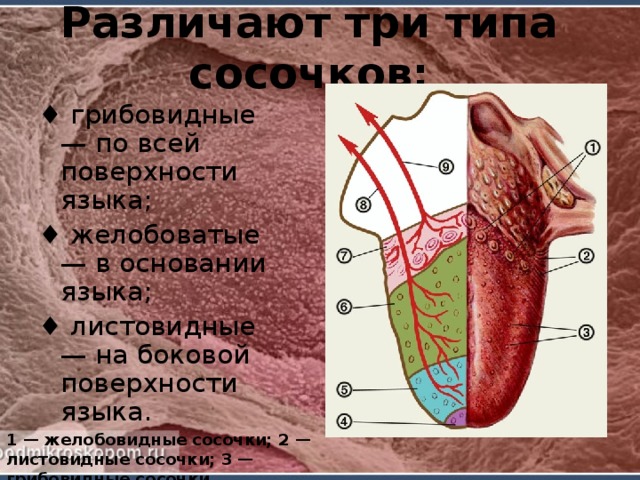 Действительно, это было доказано, идентифицировано более 20 горьких T2R. Также было показано, что каждая клетка горького вкуса экспрессирует не все горькие T2R, а только некоторые из них. 2
Действительно, это было доказано, идентифицировано более 20 горьких T2R. Также было показано, что каждая клетка горького вкуса экспрессирует не все горькие T2R, а только некоторые из них. 2
Исследования восприятия солевого вкуса показали наличие катионных каналов. По мере увеличения концентрации в ротовой полости катионы поступают в клетки солевых рецепторов, что приводит к деполяризации и, в конечном итоге, к высвобождению нейротрансмиттеров. Различные реакции на одинаковые концентрации различных соленых соединений указывают на то, что вкус соли может быть больше, чем катионные каналы на поверхности вкусовых клеток. 2
Восприятие кислого вкуса первоначально было связано с концентрацией ионов водорода. Однако с тех пор было показано, что нет прямой зависимости между pH, титруемой кислотностью и кислым вкусом. Растворы органических кислот при одном и том же рН вызывают разные реакции на кислый вкус. Точно так же растворы органических кислот одинаковой нормальности также приводят к разным реакциям на кислый вкус.